Jehová Lourenço Jr
Community Ecology - Wood Anatomy - Plant Physiology

My research
I have studied environmentally diverse tropical plant communities in Brazil (restinga habitats of the Atlantic Forest), Boreal Forests tree species in Canada, and Giant Dipterocarp tree species in Malaysia. My research focuses on using these systems to improve our understanding of plant function, adaptation to environmental change, and climatic sensitivities. My research involves intense fieldwork activities to collect samples (> 1000 trees) and measure functional traits, and I have addressed five different lines of research:
1. Drivers of species distribution and trait composition
Figure 1

SITES
floodable (○), intermediate (□), and dry (∆)


The study was conducted in Espirito Santo State - Brazil (fig. 1a), within the Paulo Cesar Vinha State park (fig. 1b), where I setup 42 plots of coastal restinga forest (fig. 2c), which is highly diverse and spans strong environmental gradients. To identify the most important environmental drivers of coastal Atlantic forest functional composition, we performed a forest inventory of all plants measuring 5 cm and above.
I collected wood samples and leaves from ~85% of the most abundant plant species and estimated height, aboveground biomass (AGB), and basal area of individual plants, and the community-weighted specific leaf area (SLA). In addition to plant traits, we measured water table depth and 25 physicochemical soil parameters. I then parameterized several differing hypotheses related to the role of nutrients and soil as they influence tropical forest diversity and functioning, represented by plant traits. Hypotheses were formalized via generalized additive models and piecewise structural equation models.
There is a strong species replacement along the water table depth gradient (fig.1c and d), which was the most important explanatory environmental variable for species distribution. The shifting along the NMDS axis (fig. 1d) is related to the taxonomic difference between plant communities, calculated by the Simpson index.
Water table depth, soil coarseness, potential acidity, sodium saturation index (SSI) and aluminum concentration were all components of the best models for AGB, height, basal area, and trait composition. Among the 25 environmental parameters measured, those related to water availability (water table depth and coarse sand), followed by potential acidity, SSI, and aluminum consistently emerged as the most important drivers of forest physiognomy and functional composition.
Increases in water table depth, coarse sand and soil concentration of aluminum negatively impacted all the measured functional traits (fig. 2 a-d), whereas SSI had a positive effect on AGB and plant height (fig. 2 a-b). These results suggest that sodium is not merely tolerated by Atlantic Forest restinga plant communities, but is important to their structure and functioning. Presence of aluminum in the soil had a complex relationship to overall basal area, possibly mediated by organic matter.
Figure 2



Simplified models obtained by piecewise structural equation models, using the forward selection analysis applied to all 25 measured environmental variables. Shown are models for the most important drivers of (a) AGB, (b) height, (c) BA and (d) SLA from 42 restinga plant communities. Sodium’s effect on AGB was marginally significant (ms; P = 0.054).
2. Using plant anatomy to predict forest diversity and sensitivity to drought
The community-weighed metrics are well-established approaches in trait-based ecology to address several ecological questions in ongoing discussions about plant hydraulics, widening the perspectives in wood anatomy survey.
The images analysis were made by the ROXAS software (fig. 1), a cutting-edge software for wood anatomy analysis, I analyzed a high diversity of wood anatomical architectures in 36 tree-species (fig. 2) to collected several wood anatomy traits (just showing four traits in fig. 3), which were calculated by the software for image analysis.
I calculated the community weighed means (fig. 3) to assess the shifting of several wood anatomy traits along the steep soil water availability gradient in the restinga habitat of the Brazilian Atlantic forest. I tested several hypotheses in ecological wood anatomy regarding to the hydraulic safety-efficiency and discussed the functionality of several wood anatomy traits.
Toward drier conditions, we found reductions in the mean and variance of traits related to the hydraulic efficiency (theoretical hydraulic conductivity [Ks], vessel-size...), whereas hydraulic safety traits increased in mean (cell-grouping and cell-density).
Further investigation of hydraulic and photosynthetic physiology of restinga plants is timely to determine how likely plants are to resist drought-stress, allowing us to predict how tropical forests will respond to the ongoing climate change (e.g. drought and warming).
Figure 1
The images were analyzed with the semi-automatic image analysis tool ROXAS v3.0 (von Arx & Carrer, 2014).
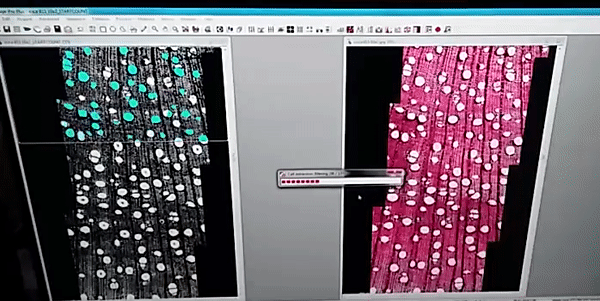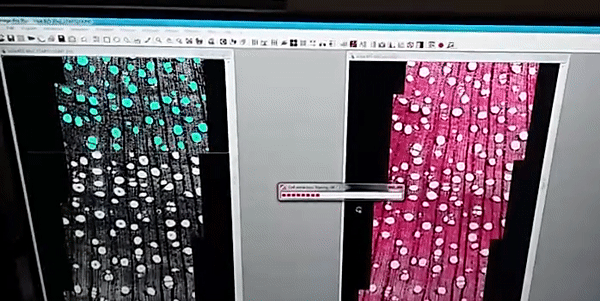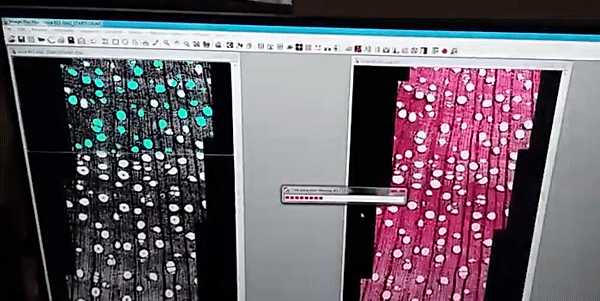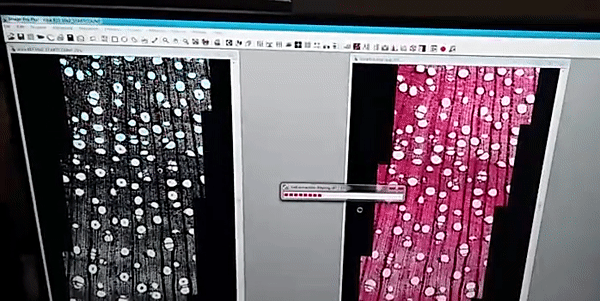
von Arx G, Carrer M. 2014. Roxas -A new tool to build centuries-long tracheid-lumen chronologies in conifers. Dendrochronologia 32: 290–293.
Figure 2
There is a high diversity of anatomical architectures in the 36 restinga tree-species collected.

Figure 3
Community weighed mean of theoretical hydraulic conductivity (Ks), cell density, vessel grouping index, and mean vessel size along the water table depth gradient. Statistically significant (*), p-value<0.05. Floodable (○), intermediate (□), and dry (∆) forest communities.

3. Climate change and plant mortality
Trait-Driver Theory or TDT (Enquist et al. 2015) proposes the investigations of the functional traits distribution by its decomposition into the first four central statistical moments - mean, variance, skewness, and kurtosis - to assess the drivers and dynamics of trait distribution in space and time.
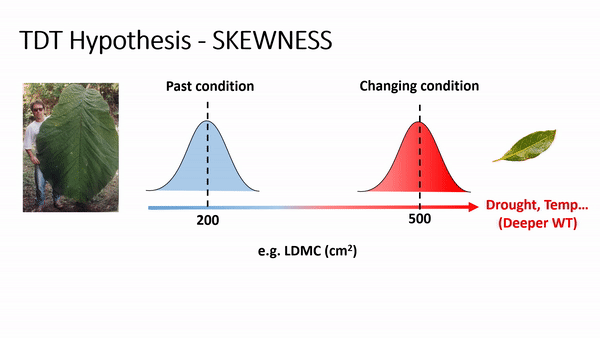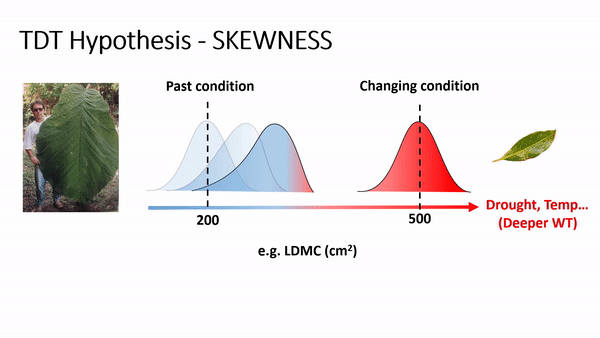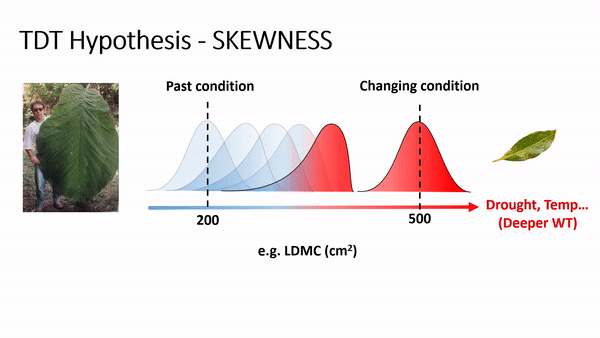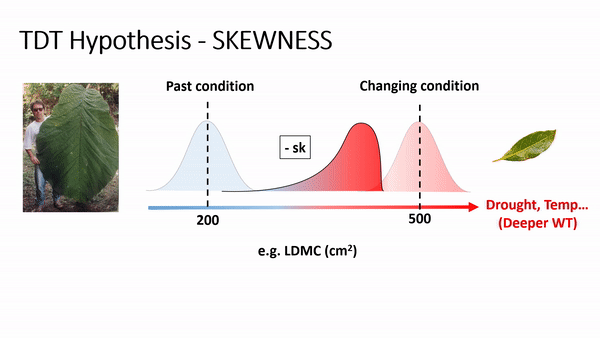
TDT predicts an increase of skewness and/or the importance of rare species or rare traits advantage in local coexistence. For example, if phenotype/environment matching is strong, then climate change (e.g., via increases in temperature and/or drought) will cause community trait distributions to track the environment due to decreasing performance or fitness of the existing dominant phenotypes and increasing performance or fitness of some currently rare phenotypes (Enquist et al., 2015, 2017). As this shift occurs, an increase in the skewness is expected, reflecting a lag in time as individuals with newly maladaptive traits slowly decline in frequency. For example, if temperature and drought increase, it is expected to have a shifting in the trait distribution of leaf dry matter content (LDMC) to the right side, producing a curve with left tail (or negatively skewed - see scheme above), as plant species with ticker leaves are more resistant to drought and warming, favoring a more conservative ecological strategy of resource use and biomass allocation in plants (Wright et al., 2004; Chave et al., 2009).

In fact, the analysis of skewness (fig. 1) for LDMC showed a consistent results to the TDT hypothesis, as the most part of the 42 communities presented negative skewness values. Similar result for wood density (WD), as denser wood are linked to smaller vessels, providing safety hydraulic conductivity (find more details in Chave et al. 2009; Hacke et al. 2001; Swenson & Zambrano 2017).
Specific leaf area (SLA) and height (H) showed opposite trends (positive skewness). Smaller leaves and plant sizes are expected to be favored in drier and warmer environmental conditions (Bennett et al. 2015, Wright et al., 2004).
Figure 1
Community kurtosis of plant height (H), leaf dry matter content (LDMC), specific leaf area (SLA), and wood density (WD). Notice the scheme beside showing normal (0), positive and negative skewed curves.


Figure 2
Frequency distribution of trees by classes of sizes (tree diameter), showing floodable, intermediate and dry communities.
The strong drought and warming occurred in the last El-nino event (2014-2016) impacted restinga plant species, with important plant mortality particularly after 2015, the driest years recorded in the past 50 years in the southeast region of Brazil (Nobre, et al. 2016). The floodable forest registered the highest amount of dead trees in 2016 (fig. 2), highlighting that species from this environment may have higher vulnerability to drought. Geonoma schottiana, Euterpe edulis, Cyathea phalerata, and Callophyllum brasiliensis are between the most affected species, with high population reductions (up to 50% for C. brasiliensis).
This experimental area was setup with the purpose of conducting a long-term experiment, which is timely to accurately predict how forest composition is expected to respond in an ongoing climate change environment.
4. Methods for assessing the 3D hydraulic architecture of trees

The assessment of the hydraulic architecture of plants may contribute to a better understanding of the structural mechanisms by which plants respond to environmental change (e.g. drought, warming...). However, this task may be challenging and time-consuming!
I have developed techniques to rapidly reconstruct the 3D hydraulic architecture of trees in high resolution via laser microscopy. This is study is part of my postdoctoral research at the
University of Quebec in Montreal (2019-2021) , and have produced two manuscripts (in review) that shed some light on the mechanisms underlying the hypothesized hydraulic safety-efficiency co-optimization in plants (Liu et al. 2019) and growth performance across soil water gradients.
5. Tip-to-base scaling of xylem hydraulics: causes and consequences


Research group preparing for one more day of fieldwork, and one of the professional tree climbers climbing a giant under the supervision of Unding.

Figure 2. Tip-to-base variation in xylem traits linked to hydraulics. Picture from Olson et al. (2020)
This research is based on wood and leaf sampling for measuring anatomical traits linked to hydraulic and physiological traits (e.g., hydraulic vulnerability curves). According to predictions and recent theories, the scaling of xylem traits, such as tip-to-base variation in vessel width, is a pervasive trait in plants selected during the evolution of terrestrial plants (West et, al, 1997; Enquist, 2002; Olson et al., 2020). Thus, the scaling of xylem traits has strong implications for how trees transport water from roots to leaves, and consequences for these plant species' climatic sensitivities (e.g., drought vulnerability).
Our preliminary results (example above by Jotan, 2024) show the expected vessel widening (basal, intermediate, and top branches), and we are still investigating how this influences water transport and drought resistance. High values of vessel widening exponent (b~3) have been found in many species, meaning that vessels are larger than expected given stem path length, i.e., giant trees present low resistance to transport water resistance but at a cost of increased drought vulnerability.

References
Bennett, A. C., Mcdowell, ... Anderson-Teixeira, K. J. Larger trees suffer most during drought in forests worldwide. Nat. Plants 1, 1–5 (2015).
Chave, J., Coomes, D., Jansen, S., Lewis, S. L., Swenson, N. G., & Zanne, A. E. (2009). Towards a worldwide wood economics spectrum. Ecology Letters, 12(4), 351–366. doi:10.1111/j.1461-0248.2009.01285.x
Enquist BJ. (2002). Universal scaling in tree and vascular plant allometry: toward a general quantitative theory linking plant form and function from cells to ecosystems. Tree Physiology 22: 1045–1064.
Enquist, B. J., Bentley, L. P., Shenkin, … Malhi, Y. (2017). Assessing trait-based scaling theory in tropical forests spanning a broad temperature gradient. Global Ecology and Biogeography, 26(12), 1357–1373. doi:10.1111/geb.12645
Enquist, B. J., Norberg, J., Bonser, S. P., Violle, … Savage, V. M. (2015). Scaling from Traits to Ecosystems: Developing a General Trait Driver Theory via Integrating Trait-Based and Metabolic Scaling Theories. Advances in Ecological Research (1st ed., Vol. 52). Elsevier Ltd. doi:10.1016/bs.aecr.2015.02.001
Hacke, U. G., Sperry, J. S., Pockman, W. T., Davis, S. D., & McCulloh, K. A. (2001). Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia, 126(4), 457–461. doi:10.1007/s004420100628
Liu H, Ye Q, Gleason SM, He P, Yin D. 2020. Weak tradeoff between xylem hydraulic efficiency and safety: climatic seasonality matters. New Phytologist 229: 1440–1452.
Marengo, J. A., Alves, L. M., ... Moraes, O. L. L. (2017). Climatic characteristics of the 2010-2016 drought in the semiarid Northeast Brazil region. Anais Da Academia Brasileira de Ciências, (0), 1–13. doi:10.1590/0001-3765201720170206
Olson ME, Anfodillo T, Gleason SM, McCulloh KA. (2020). Tip-to-base xylem conduit widening as an adaptation: causes, consequences, and empirical priorities. New Phytologist 229: 1–17.
Swenson, N. G., & Zambrano, J. (2017). Why wood density varies across communities. Journal of Vegetation Science, 28(1), 4–6. doi:10.1111/jvs.12510
West GB, West GB, Brown JH, Enquist BJ. (1997). A general model for the origin of allometric scaling laws in biology. Science 276: 122–126.
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., … Villar, R. (2004). The worldwide leaf economics spectrum. Nature, 428(6985), 821–827. doi:10.1038/nature02403